2.大腸菌のポリアミン代謝系〜ポリアミン代謝とグルタチオン代謝の関係
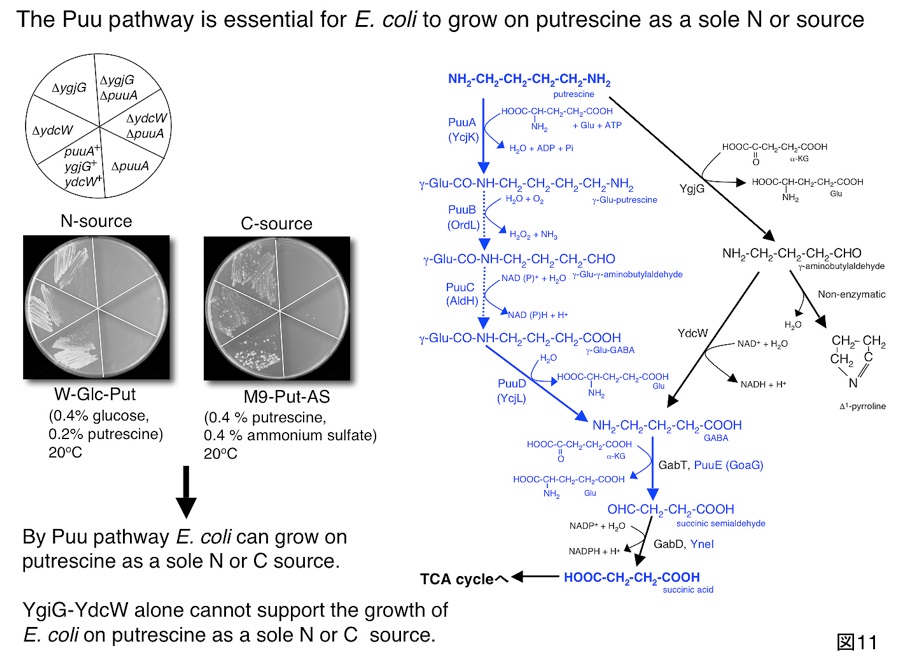
図9はpuuA遺伝子の転写に関するreal-time RT PCRの結果を示しています。
プトレッシンが培地中に存在するとき、PuuAの転写は著しく活性化され、またPuuA遺伝子の転写がPuuRの存在により制御されていることが分かります。
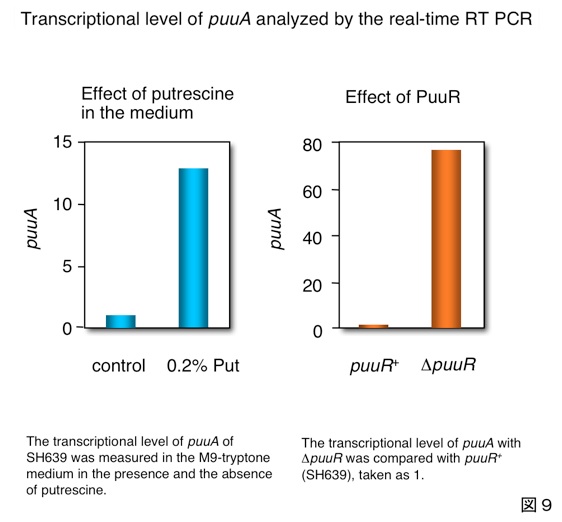
次にγ-グルタミル結合を切断するPuuDです。図10に示すようにオペロンの構造からPuuDをノックアウトすることはできません。そこで、活性を失うC114A変異株を使ったところ、γ-グルタミル-GABAが蓄積することが分かりました。
PuuAがせっかくプトレッシンをγ-グルタミル化しても、そのはしからPuuDがγ-グルタミル結合を切断してしまっては意味がありませんが、PuuDを精製して調べたところ、γ-グルタミルプトレッシンよりγ-グルタミル-GABAに対するアフィニティーが高く、また加水分解活性も高いことが分かり、PuuDは主としてγ-グルタミル-GABAを基質としていることが分かりました。
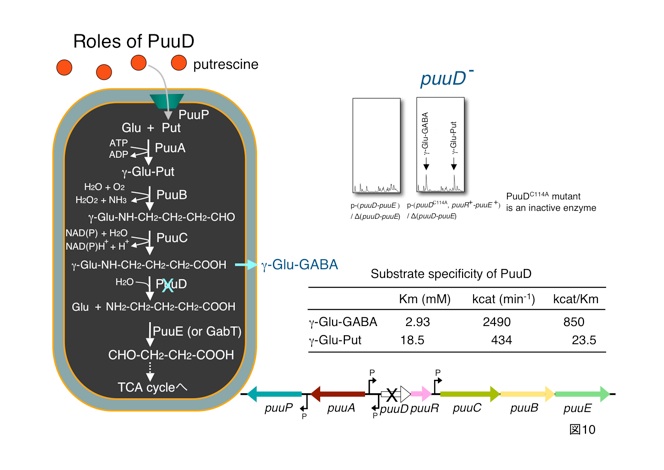
プトレッシン異化経路として大腸菌には、私たちが発見したPuu代謝系(図11に青色で表示)のほかに、昔から知られているYgjG-YdcW代謝系があります。
遺伝子破壊株はプトレッシンを唯一N源あるいはC源とする寒天培地に生育できません。一方、YgjGあるいはYdcW遺伝子破壊株はいずれのプレートにも生育できます。つまり、Puu代謝系は大腸菌がプトレッシンを唯一N源あるいはC源として生育するのに必須ですが、 YgjG-YdcW代謝系はそうではないことが分かりました。