
安定同位体の解説
植物の安定同位体についての解説と利用方法です。安定同位体を利用した研究については、総合地球環境研究所から出されている「同位体環境学がえがく世界 2023」や、2009年に北海道大学低温研究所発行の「光合成研究法(低温科学)」、日本生態学会誌などに日本語でのレビューや解説が出されているので、そちらも参考にして下さい。ここでの解説は「同位体環境学がえがく世界」、「光合成研究法」と2006年度春学期に関西大学で行った講義(自然環境学研究B)をもとにしています。
安定同位体を使った研究例1:大気汚染の街路樹への影響を評価する
1. はじめに
日本の都市部で、大気汚染物質である窒素酸化物の問題が最も顕著だったのは1985年前後でした。1990年代なかば以降は、日本の大気中の窒素酸化物の量は着実に減少しています。一方、2000年代以降は、アジア地域で中国やインドなどの著しい経済成長に伴って大気汚染が劇的に悪化し、大気汚染の問題は、アジア地域における最も重要な環境問題の一つとなっています。都市における窒素酸化物の主な排出源は、自動車です。日本の都市部で大気中の二酸化窒素量が着実に減少してきた背景には、排ガス浄化装置の搭載など、トラックの環境性能が大幅に向上したことがあります。しかし、交通量が非常に多い幹線道路沿いなど、京都市の二酸化窒素のレベルが高い地点では、2019年時点でも1年の半分以上の日数で環境基準の濃度(0.02ppm)を超えています。大気汚染は、日本においても過去の問題とはいえないのです。
2. 炭素安定同位体比から街路樹の大気汚染ストレスを診断する
都市に植栽されている街路樹は、樹冠で大気汚染物質を吸収・捕捉したり、木陰をつくって高温化を抑えたり、光合成で二酸化炭素を吸収するなどの多くの効用を持っています。光合成は街路樹の生長や生存を支える生理的な働きであるため、光合成活性を維持することは、都市部で樹木が生存していくためには必須です。しかし、光合成に必要な二酸化炭素を取り込む「気孔」は、大気汚染ストレスを感じるといち早く閉じてしまい、その結果、光合成が低下してしまいます。
植物の葉に含まれる光合成産物の炭素安定同位体比(δ13Cplant)は、植物の光合成活動による同位体比の変化(Δ13C)と、大気の同位体比(δ13Cair)によって決まります。Δ13Cは、平均的な気孔の開度を反映します(図1)。気孔があまり開いていないと、軽い方の同位体(12C)が植物に取り込まれにくくなるためです。ストレスを感じると、多くの場合、気孔は閉じていきます。このようなことから、Δ13Cの値から、植物が受けている大気汚染ストレスを診断できます。
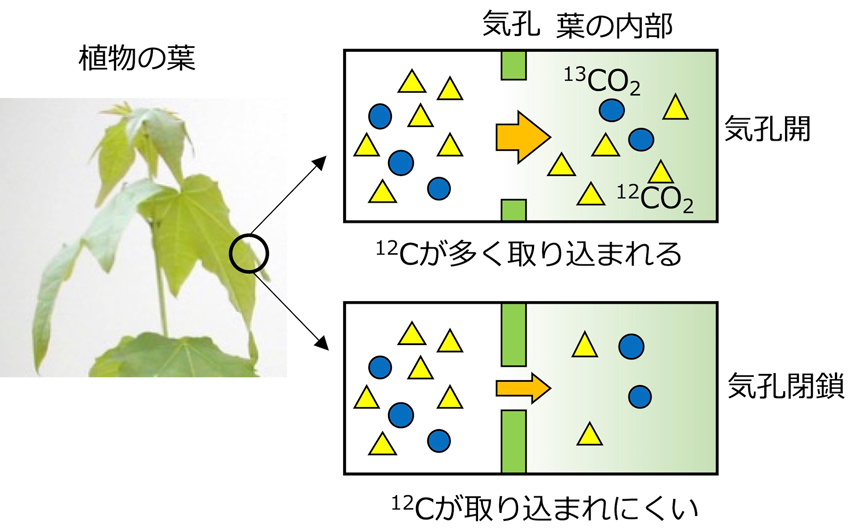
【図1】葉の内部への炭素安定同位体12Cの取り込まれ方は、気孔が開いているときと閉じているときで異なる。
木本植物の生理生態(2020)を改変。
3. 京都市内における交通量の違いと街路樹の大気汚染ストレス:Δ13Cの空間的変化
わたしたちはΔ13Cを使って、京都市内の場所による交通量の違いが、街路樹の大気汚染ストレスとどのように関係しているのかを調査しました。その結果、街路樹であるヒラドツツジは、交通量が多い、つまり二酸化窒素量が多い場所にあるほど、気孔が閉じていました。このことから、大気中の二酸化窒素が、ヒラドツツジの光合成機能に悪影響を及ぼしていることが分かりました(図2)。
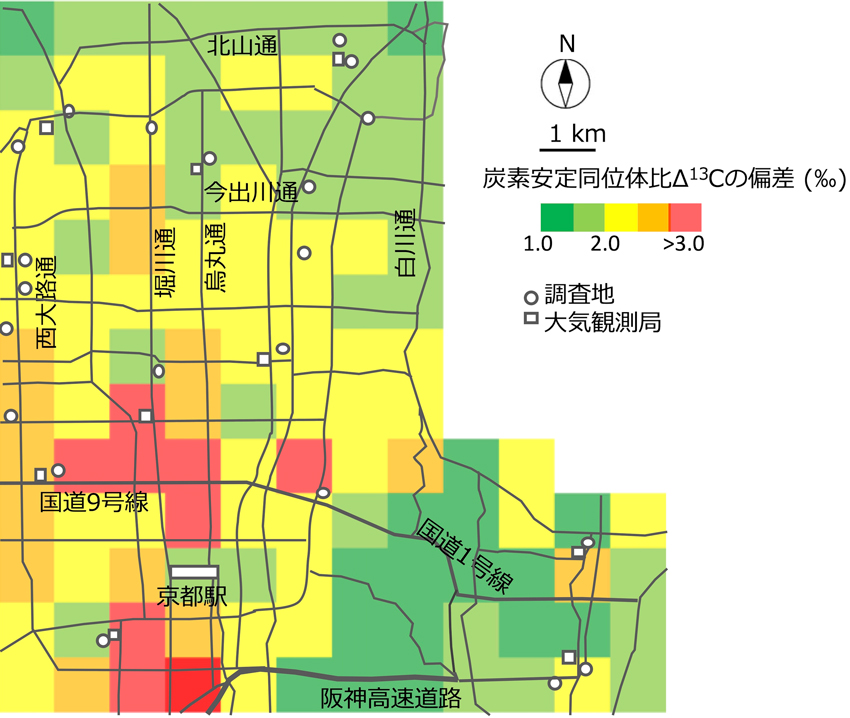
【図2】常緑低木であるヒラドツツジの葉のΔ13Cの偏差を京都市内でマッピング。
Δ13Cの偏差が大きいほど強いストレスがかかっている。
4. 街路樹の炭素安定同位体比からCOVID19による産業活動停滞の影響を検出する:Δ13Cの時間的変化
大気汚染の大きな要因になっているのは、トラックから排出される窒素酸化物です。2020年から始まった新型コロナウイルス感染症(COVID19)の流行は、世界的な経済産業活動の低下を引き起こし、日本も例外ではありませんでした。この経済産業活動の低下により、トラックなどの車両の通行台数が減り、大気中の二酸化窒素量も減ったと考えられます。その結果として街路樹では、光合成機能低下が緩和された可能性があります。わたしたちは2005年から2023年にかけて断続的に、京都市を中心とした都市域で街路樹の葉を採取してΔ13C解析を行っています。葉のΔ13Cの経年変化を追跡することで、街路樹の光合成機能に、COVID19による経済産業活動の低下がどのように影響したのかを検出できる可能性があります。
5. COVID19は街路樹の光合成機能に影響せず
京都市の2005年−2023年の二酸化窒素濃度を解析したところ、特に交通量が多い大気汚染物質観測局で、リーマンショックが起こった2008年から2009年に大幅な落ち込みが見られ、さらに2020~2023年にも落ち込みがみられました(図3)。2020~2023年に、COVID19による影響が二酸化窒素濃度の減少となって現れている可能性があります。
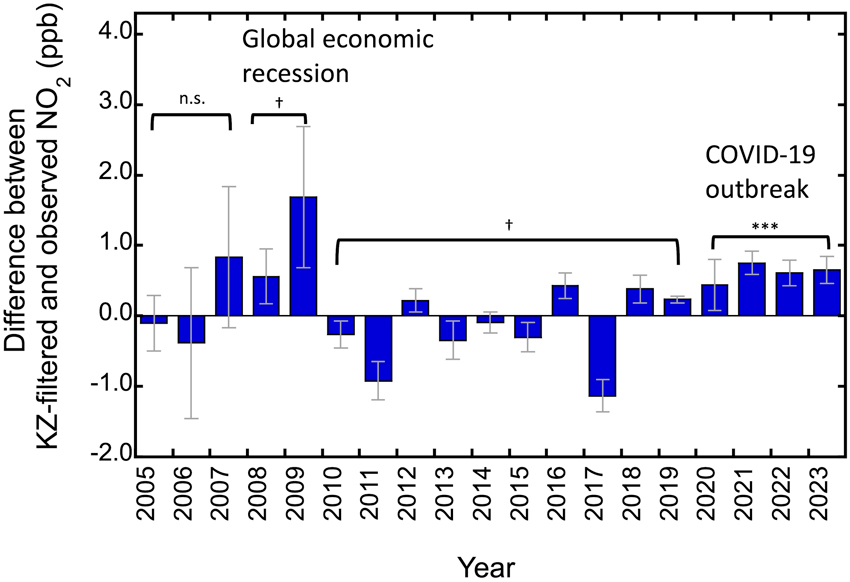
【図3】2007-2019年のデータの直線回帰によって予測されるNO2濃度と、実測値との偏差。
値が大きいほど、NO2が予測値より減少していることを示す。
NO2データ:http://www.nies.go.jp/igreen/td_down.html
2005年-2019年のΔ13Cデータを、COVID19の影響があった2020~2023年データと比較してみました。イチョウ、ヒラドツツジともに、COVID19の影響がなかったと仮定したときの値と実測値との間に、有意な差はありませんでした。このことから、どちらの種でも、COVID19の影響は安定同位体比に現れていない、すなわち、COVID19によりストレスが緩和されたことはなかったといえます。光合成測定についても同様に、COVID19によりストレスが緩和され、光合成が増加した、という結果は、どちらの種でも得られませんでした(図4)。
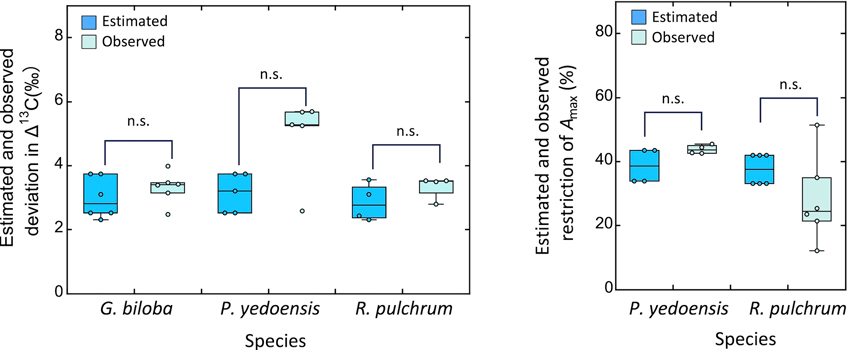
【図4】2007-2019年のデータの直線回帰によって予測されるΔ13C/光合成速度と、実測値。
予測値と実測値の差が大きいほど、COVID19の影響が大きいことを示す。
どちらの種でも、予測値と実測値の間に有意差はない。
皮肉なことではあるのですが、COVID19の影響により世界の多くの都市で経済産業活動が停滞し、その結果、大気汚染は(一時的に)緩和されています。しかし、日本においては、すでに大気汚染対策が進んでいたこともあり、COVID19の影響によるNO2濃度の減少は顕著ではなく、したがって、街路樹へのストレスが緩和されることもありませんでした。
街路樹に対して何がストレスになるか、を考えると、日本においては大気汚染よりも、「長い夏(高温が長く継続する)」の影響が大きくなる可能性があります。安定同位体は、このような中長期的な気候変動に対する植物の応答を評価する、有用なツールと言えます。
安定同位体を使った研究例2:葉肉コンダクタンスの推定
葉肉コンダクタンスの推定方法については、こちらのマニュアルを参考にしてください。
炭素安定同位体を利用して植物の葉内のCO2拡散コンダクタンスを測定する方法について解説します。炭素安定同位体については炭素安定同位体の章を参照して下さい。
1. 測定システム
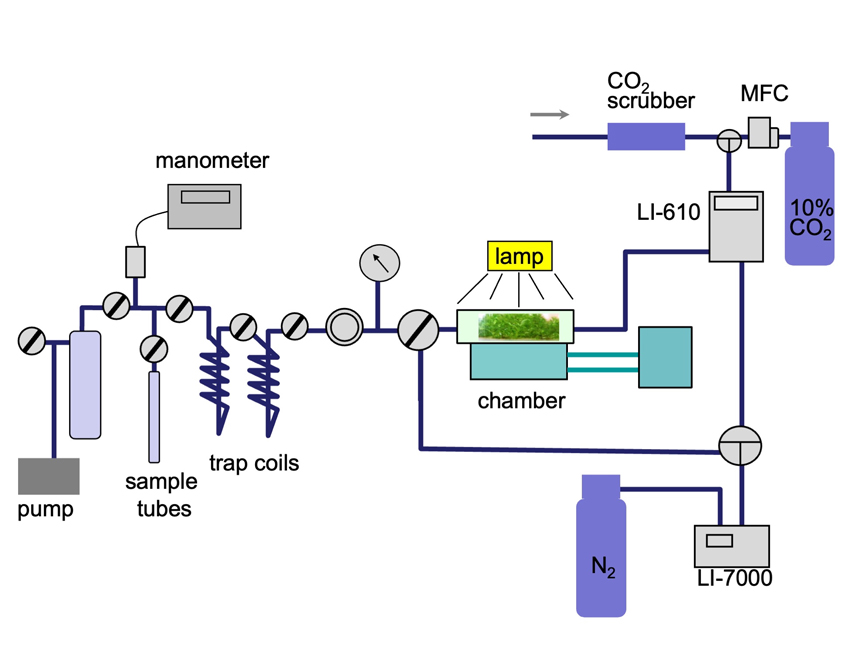
測定システムは、メーカーから購入した既製品の測定装置に、特注品としてメーカーに製作を依頼したいくつかのパーツを組み合わせて製作しました。光合成測定(右側)とCO2収集(左側)の2つのパートに分かれています。
(1) 光合成測定装置の概略
光合成測定装置を自作するにあたっては、各パーツの材質を考慮することと、測定機器を信頼できるメーカーから購入することが重要です。細かいノウハウについては Pearcy et al. (ed.) Plant Physiological Ecology (filed methods and instrumentation) 2000, Kluwer Academic Publishers に詳しく掲載されています。光合成を測定するためには、葉をチャンバに挟んでCO2吸収量と蒸散量を赤外線ガス分析計(LI6162, Li-Cor)で測定します。葉に送る空気の量や湿度、また葉の温度を適当な値に制御します。
■空気の流量とCO2濃度および湿度調整:屋外から大気を露点発生器(Li-610, Li-Cor)を介して吸引しソーダライム(およそ800g)を通してCO2を取り除きます。CO2フリーの空気に、10%のCO2を含んだガスボンベから供給されるCO2を混合し、マスフローコントローラー(CR700, KOFLOK)でCO2濃度と流量を調節してチャンバに供給します。空気中の水蒸気量は露点発生器(Li-610, Li-Cor)で調節します。
■チャンバの温度調整:恒温循環槽で温度調整した水をチャンバ下部の水槽に通すことによって調整します。
■光源:高輝度LEDランプ(LEDRB-630DL,オプトコード)を使用しています。
■葉温の測定:熱電対の一方を氷水中にセットし、もう一方を葉の表面に触れるようにセットします。
■測定値のモニター:空気の流量や露点、葉温、赤外線ガス分析計で測定したCO2量と水蒸気量はすべてデジタル式の記録計(MV100、横河電機)でモニターします。
(2) 収集装置の概略
左側のCO2収集装置(真空ライン)は、パイレックスガラス製の特注品です(光信理化学製作所に製作を依頼)。真空ポンプ(GLD-050, Sinku Kikou)を常時稼働させます。ラインの真空度はピラニゲージ(model 937, MKS instruments)でモニターします。
(3) 光合成・拡散コンダクタンス測定の手順
上記の測定装置を使った光合成・拡散コンダクタンス測定の手順マニュアルは ここからダウンロードしてください。
上記の測定装置を使った光合成・拡散コンダクタンスの計算方法マニュアルは ここからダウンロードしてください。
安定同位体の基礎―安定同位体を使う意義
安定同位体は環境に対する植物の反応を研究するために利用される研究手法である。植物の環境反応研究は研究対象の空間的なスケールで分類することができ、大きな方から生態系、群落、個体、組織、細胞、オルガネラとなる。おおよそのスケールはもっとも大きな生態系は106m、最も小さいオルガネラは10-6mである。研究手法は対象によって様々であり、オルガネラや細胞では遺伝子やタンパク解析など分子・生化学的手法が、葉や個体ではガス交換法などの生理的手法が、群落や生態系では微気象的方法やリモートセンシングなどが用いられる。安定同位体は他の方法と比べると、細胞レベルから生態系レベルまで広範囲に利用できること、また積算的な情報が得られ比較的分析コストが安価であるという特色がある。
植物の環境研究に使われる手法
| 方法 | 測定スケール | 利点 | 欠点 |
|---|---|---|---|
| 安定同位体 | 細胞から生態系まで | 積算的な情報が得られる 分析が容易でコストも安い |
影響因子の特定が難しい場合がある |
| 生化学的方法 | 細胞 葉の切片 植物抽出物(タンパク質など) |
詳細な分析が可能で反応機構の研究に適している | 生体外測定のため生体内での再現性が不明確 |
| 生理学的方法 (ガス交換法) |
葉 シュート |
生きた植物で測定できる 測定が比較的簡便 |
測定時の環境条件に大きく影響される 瞬間的な情報しか得られない |
| 生態学・微気象的方法、リモートセンシング | 群落、生態系 | 大規模スケールで推定可能 | 時間とコストがかかる 植物の反応との関連づけが難しい |
安定同位体について
同位体とは、ある特定の元素に属し、核の中性子数が異なるため質量が異なっている原子のことをいう。同じ元素に属する同位体は原子量は異なるが原子番号は同じであり、プラスの電荷もマイナスの電荷も持たない。同位体は、時間の経過とともにエネルギーを放射して別の元素に壊変する放射性同位体と、時間が経過しても不変である安定同位体 stable isotopeとに分けられる。放射性同位体の安定性は半減期(half-life)であらわされる。半減期とはある系に存在する放射性同位体の数が半分になるのに要する時間である。半減期には同じ元素の同位体の中でも十数秒(10C)から数千年(14C)までの幅がある。一方、安定同位体の量は存在比 abundanceであらわされる。ある特定の元素の中では1つの安定同位体の存在比が飛び抜けて高く、例えば12Cは全てのCの98.9%を占める。
| 元素 | 安定同位体 | 存在比(%) | 放射性同位体 | 半減期 |
|---|---|---|---|---|
| 水素 | 1H 2H |
99.985 0.015 |
3H | 12.32y |
| 炭素 | 12C 13C |
98.89 1.11 |
10C 11C 14C 15C |
19.3s 20.3m 5715y 2.45s |
| 酸素 | 16O 17O 18O |
99.76 0.04 0.20 |
14O 15O |
70.6s 122.2s |
| 窒素 | 14N 15N |
99.63 0.37 |
12N 13N |
0.1s 10m |
| 硫黄 | 32S 33S 34S 36S |
95.0 0.76 4.22 0.014 |
35S | 87.51d |
Nilsen & Orcutt (1996) Plants under Stress. John Wiley & Sons., Pearcy et al. (2000) Plant Physiological Ecology. Kluwer.
サンプルの安定同位体比は標準物質に対する偏差であらわされる。この偏差は非常に小さいので1000分偏差(パーミル:‰)であらわされることになっている。ある物質 X について、2種の安定同位体の比率をRとすると、安定同位体比は次の式で計算される。
δX ={Rサンプル/R標準物質- 1 }× 1000(‰)
| 元素 | 安定同位体の比率 R | 表記 | 測定気体 | 測定精度(‰) |
|---|---|---|---|---|
| H | 2H/1H (D/H) | δD | H2 | 0.2* |
| C | 13C/12C | δ13C | CO2 | 0.01** |
| O | 18O/16O | δ18O | CO2 | 0.01** |
| N | 15N/14N | δ15N | N2 | 0.02* |
| S | 34S/32S | δ34S | SO2 | 0.5*** |
*200µL **100µL ***50µg
炭素安定同位体比
炭素の安定同位体のほとんどは12Cで、13Cは1%程度しか含まれていない。12Cと13Cは、大気中や水中を拡散する速度や化学反応の反応性が異なる。軽い同位体12Cの方が拡散する速度は速く、一般的には化学反応に対する反応性も高い。有機物質の同位体組成はいったん乾燥させればほとんど変化しないため、分解が起こらない状態で保存されているサンプルの炭素同位体組成は長期間一定である。
植物は、気孔からCO2を取り込んで炭素化合物を合成する。大気中のCO2から炭素化合物がつくられるまでにはさまざまな化学反応プロセスがある。そのため、大気中CO2の炭素と植物に含まれる炭素化合物の炭素の重さを比べると、植物の炭素化合物の炭素の方が少しだけ「重い」安定同位体が少なくなる。このように同位体の比率が変わることを、同位体分別(どういたいぶんべつ)とよぶ。(注:ふんべつ とは読まない)
炭素の安定同位体比は、国際的に定められた標準物質 PDB(Pee Dee belemnite:アメリカ・サウスカロライナ州にあるPee Dee層から産出したイカの仲間 belemniteの化石に含まれる炭酸カルシウム)に対する13C/12Cの偏差としてあらわされる。13C/12CをRとすると、炭素安定同位体比δ13Cは次のように定義される。
δ13C(‰)=(Rサンプル/R標準物質 - 1)×1000;
ほとんどの物質では、炭素の同位体には「重い」同位体13Cが標準物質よりも少ししか含まれていないため、同位体比としてはマイナスの値となる。例えば、大気中のCO2は -8‰ であるが、これは大気中のCO2が標準物質よりも 8/1000 だけ13C/12Cが少ないことを示している。
炭素安定同位体比の決定要因
植物全体の炭素安定同位体比は主に次のような4つの要因で決まる。
- 大気中のCO2の炭素安定同位体比
- 光合成による同位体分別
- 炭素化合物の組成
- 転流
大気CO2の炭素安定同位体比
| 測定場所 | 炭素安定同位体比(‰) |
|---|---|
| 上層大気 | -8 |
| 都市部の大気* | -11.6 |
| 山間部の大気** | -10 京都市美山町国道 -12 タイ・ナラチワ熱帯林 -9 夏の北海道落葉広葉樹林 |
| CO2ボンベ | -24 ~ -38 |
自動車の排気ガスのように化石燃料から発生するCO2は、上空の大気に比べると「重い」安定同位体がずっと少なくなる。つまり、都市部や工業地帯の大気中のCO2は、山間部や砂漠のような人間の活動の影響が少ないところとは、炭素の安定同位体比が異なる。
また、森林の大気のCO2には土壌呼吸によってでてくるCO2や、植物の呼吸によってでてくるCO2が混合している。呼吸によってでてくるCO2には「重い」安定同位体が少なくなっている。そのため、呼吸ででてきたCO2がたまりやすい条件―例えば地表付近とか、樹木の密度が高い森林(熱帯林など)、あるいは夜間―であるほど、森林の大気のCO2にはさらに「重い」安定同位体が少なくなる。
光合成による同位体分別
| 過程 | 分別(‰) |
|---|---|
| CO2の水への溶解 | 1.1 |
| CO2の水和 | -9.0 |
| 大気中でのCO2拡散 | 4.4 |
| 水中でのCO2拡散 | 0.7 |
| カーボニックアンヒドラーゼ触媒によるCO2の水和 | 1.1 |
| PEPカルボキシラーゼによるカルボキシル化 | 2.0 |
| Rubiscoによるカルボキシル化 | 29.0 |
光合成では大きな同位体分別がおこることが知られている。その主なプロセスは
- CO2が気孔から取り込まれるとき
- 酵素によってCO2が同化されるとき
の2つに分けることができる。光合成によってCO2が固定されるときにはたらく酵素は植物が利用している光合成経路によって異なり、C3植物では主にRubisco、C4植物とCAM植物では主にPEPカルボキシラーゼとRubiscoである。Rubiscoによる同位体分別は 29‰程度であるのに対して、PEPカルボキシラーゼによる同位体分別はずっと小さく、2‰ 程度であると考えられている。 一方、CO2が気孔から取り込まれるときにおこる同位体分別は 4.4‰とされている。
おおきな同位体分別を示す酵素Rubiscoが主にはたらくC3植物は、C4植物やCAM植物と比べて「重い」安定同位体が少なくなっている。C3植物の炭素の安定同位体比は平均値で -28‰ であるが、C4植物では -14‰である。CAM植物の値は-10 ~-20‰であり、昼間と夜間に固定されるCO2の比率によっても変わる。
光合成の際に起こる同位体分別は、記号Δ(ギリシャ文字の大文字のデルタ)で表される。C3植物では、もっとも簡略的にあらわすと次のようになる(Farquhar et al. 1982)。
Δ = 4.4 [ ( Ca- Ci) / Ca ] + b Ci/Ca(‰)
CaとCiは、それぞれ大気と葉内(細胞間隙)のCO2分圧をあらわしている。4.4は、CO2が気孔から取り込まれる際に、12CO2と13CO2の拡散速度の違いによって生じる分別(‰)である。bは、CO2固定酵素によってCO2が同化されるときの分別で、27‰と考えられている。この式から、C3植物の同位体分別は、酵素反応によってほとんど決まることが分かる。
CO2分圧と同位体分別との関係は、次のように説明することができる。Rubiscoは12CO2の方とよく反応するため、Rubiscoの近くの空気(細胞間隙)では13CO2よりも12CO2が多く消費され、その結果相対的に13CO2が増える。気孔が閉じてくると細胞間隙での13CO2の濃度が高くなっていくため、Rubiscoは13CO2とも反応するようになってくる。一方、気孔が十分に開いていれば、細胞間隙と大気のCO2は十分に混合されるので、細胞間隙での13CO2濃度は大気よりもそれほど高くならない。このときにはRubiscoが13CO2と反応する割合は相対的に低くなる。このようなことから、大気中のCO2分圧 Ca が一定である場合には、C3植物の同位体分別はCiが小さいほど小さくなる(相対的に軽い同位体が少なくなる)ことになる。
なお、C4植物では、CO2はまずPEPカルボキシラーゼによって葉肉細胞で固定された後、維管束鞘細胞でRubiscoによって再固定される。従って、CO2分圧との関係をC3植物のような簡略化された式で表すことはできない。しかし、細胞間隙のCO2分圧の変化に対する同位体比の変化はC3植物よりもずっと小さいことが分かっている。また、維管束鞘細胞で固定されるCO2の比率が高いほど同位体比は低くなる(よりC3植物に近い値になる)ことが知られている。(Farquhar et al. 1982,Buchmann et al. 1996)。
炭素安定同位体比の利用方法
水利用効率
炭素の安定同位体比は、植物の水利用効率に対する指標として、欧米の研究者を中心に広く利用されている。水利用効率(water use efficiency, WUE)は蒸散(E)と光合成速度(A)の比で表される値で、葉から失われた水に対してどの程度炭素を固定できたかを表している。おおざっぱにいうと、水利用効率は、植物に乾燥ストレスがかかっているほど高くなる。水利用効率 WUE は、植物葉内のCO2分圧(Ci)と植物体周辺のCO2分圧(Ca)及び葉-大気間の水蒸気圧差(VPD)と次のような関係にある。
WUE = A/E = Ca(1 - Ci/Ca) / (1.6 VPD) (4)
炭素の安定同位体比も、水利用効率と同じようにCi/Caと直線的な相関があるため、炭素の安定同位体比から水利用効率を推定することができる。炭素の安定同位体比がよりプラスの値であるほど水利用効率は大きいということになる。例えば、山の斜面に生えている樹木の炭素安定同位体比を調べて、上部の樹木が -27‰であるのに対して下部の樹木が -30‰ であったとすると、斜面上部の樹木の方が水利用効率が大きい、つまり、より乾燥ストレスがかかっているらしい、と解釈できる。
野外の植物の水利用効率は、携帯型の光合成・蒸散測定措置(Li-Cor 6400など)で実測することができる。あえて炭素の安定同位体比を水利用効率の指標として使う意義は、
- 生育期間の積算的な水利用効率を反映していること
- 乾燥した植物標本でも測定できること
- 多数のサンプルを短時間で測定できること
- 年輪の解析では、比較的長期間の経時変化を追跡できる
つまり、測定装置を持ち込むのが難しい場合(樹木の最上部や高山など)や、比較的大きなスケールでの仕事に威力を発揮するといえる。大きなスケールの仕事は国内ではまだあまり例がないが、オーストラリアで調べられた例では、樹木の群落レベルでの安定同位体比の変化を海岸から内陸まで数百キロメートルにわたって調べたところ、降水量が少ないほど群落レベルで葉の水利用効率が高くなることが示唆されている(Stewart et al. 1995)。
わたしたちが行った仕事としては、林床の植物と上層の樹木の間では水をめぐる競争が起きているため、管理されている(下草を刈っている)アカマツ林の方が乾燥ストレスがかかりにくいことを示した例がある(Kume et al. 2002)。林の管理が樹木の水利用や成長に大きな影響を及ぼすことは、北米のポンデローサ松でも示されている(McDowell et al. 2003)。
C3植物とC4植物を分ける
| 科 (単子葉類) |
トチカガミ | カヤツリグサ | イネ | ||||
|---|---|---|---|---|---|---|---|
| 種数 | >1 | 1330 | 4600 | ||||
| 科 (双子葉類) |
フウチョウソウ | アカザ | オシロイバナ | キク | キツネノマゴ | ゴマノハグサ | ザクロソウ |
| 種数 | >2 | 550 | >5 | 150 | 80 | 14 | >4 |
| 科 (双子葉類) |
スベリヒユ | タデ | トウダイグサ | ナデシコ | ハマビシ | ハマミズキ | ヒユ | ムラサキ |
|---|---|---|---|---|---|---|---|---|
| 種数 | 70 | 80 | 250 | 50 | ~50 | ~30 | ~250 | >6 |
C3植物とC4植物は、炭素の安定同位体比が異なる。このことを利用すると、植物の炭素安定同位体比から光合成回路を推定することができる。現在のところC4植物が含まれることが明らかになっている科は表の通りである。応用として、土壌の炭素の安定同位体比から、大規模な気候変化(乾燥化)に伴って植生中のC3植物とC4植物の割合がどのように変遷したのか(Schwartz et al. 1996)を推定できる。C4植物を含む最大の科であるイネ科の Alleteropsis属、Neurachne属、Panicum属 の中にはC3-C4中間型の性質をもつ種があるため、炭素安定同位体比からでのみ光合成型を決めることはできない。また、CAM植物とC3あるいはC4植物も、炭素安定同位体比によって区別することはできない。
葉内CO2拡散抵抗の推定
炭素の安定同位体分析とガス交換法による光合成測定とを組み合わせると、葉内CO2拡散抵抗の推定ができる。
森林のCO2・H2O循環モデルへの応用
大気や樹木の葉の炭素の安定同位体比は、森林のCO2・H2O循環モデルに組み込んで利用されている。森林の CO2・H2O循環は地球環境問題と関連する重要なテーマで、最近も多くの研究がなされている(Kosugi et al. 2003、Baldocchi & Bowling 2003など)。
窒素安定同位体比
窒素には2つの安定同位体14Nと15Nがある。14Nは存在比が高く大気中の窒素の99.633%を占める。大気の窒素の同位体組成は一定であるため(JunkとSuek 1958、Mariotti 1983)、大気中の窒素は窒素安定同位体の標準物質として使われている。すなわち、大気中窒素の安定同位体比は0‰である。14N/15NをRとすると、窒素安定同位体比δ15Nは次のように定義される。
δ15N(‰)=(Rサンプル/R標準物質 - 1)×1000;
自然界にある植物の窒素安定同位体比は-5~+10‰である。
1. 窒素安定同位体比の決定要因
| 過程 | 同位体分別(‰) | |
|---|---|---|
| 非生物的過程 | ||
| 水中でのNO3-、NH3、NH4+の拡散 | 1.0 | |
| 大気中でのNO3-、NH3、NH4+の拡散 | 1.8 | |
| 土壌中の生物的過程 | ||
| N2O還元 | 3.4~3.9 | |
| 脱窒 | 2.8~3.3 | |
| 窒素固定 | -0.9~4.1 | |
| 硝化 | 1.5~3.5 | |
| 植物 | ||
| アンモニア同化 | 0.9~2 | |
| 硝酸同化 | 0.3~3 | |
炭素の安定同位体と同じように、植物の窒素安定同位体は次のような要因に影響を受ける。
- 窒素供給源の窒素安定同位体比
- 窒素代謝過程での同位体分別
- 窒素化合物の組成
- 転流
植物への窒素供給源は、窒素固定をしない植物では土壌窒素、窒素固定する植物では土壌と大気中の窒素である。土壌表面の窒素は大気と比較すると15Nが多く、窒素安定同位体比の平均値はおよそ9.2‰である。一般的には、土壌が深くなるほど15Nが増加し、安定同位体の値はよりプラスになる。
炭素の場合と同様、窒素についてもほとんどの反応では重い同位体15Nが分別される。ただし、窒素固定反応では軽い同位体14Nが分別される場合がある。そのため、炭素の場合とは異なり、植物の窒素安定同位体比はプラスの値もマイナスの値もとりうる。
植物によって利用される土壌窒素は、硝酸態(NO3-)かアンモニア態(NH4+)となっている。アンモニア態あるいは硝酸態で窒素が吸収される場合、最大3‰の分別が生じる。また、窒素固定を行う植物の場合、植物組織の中では根粒に15Nが濃縮されて他の組織よりも窒素安定同位体比が高くなる。炭素と比べると窒素の同位体分別は小さく、また同位体分別がよく分かっていない反応過程も多いため、窒素の安定同位体比と植物の生理的な過程とを関連づけるのは難しい場合が多い。
窒素安定同位体比の利用方法
窒素安定同位体比がもっともよく利用されているのは、植物による窒素固定の研究である。窒素の同位体分別は小さいため、植物の窒素安定同位体比は窒素供給源の安定同位体比を反映すると考えてよい。窒素固定をする植物の窒素供給源は土壌と大気の2つである。土壌表面の窒素は平均9.2‰であり、大気中窒素の安定同位体比は0‰である。したがって、窒素固定を行って大気から窒素の供給を受けているマメ科の植物は、窒素固定をしない植物よりも安定同位体比が低くなる傾向を示す。
植物体の窒素安定同位体比の測定方法は、炭素安定同位体比の測定方法に準ずる。
4. 水素・酸素安定同位体比
植物の水素および酸素の安定同位体比は、植物による水利用と深い関係がある。安定同位体比の標準物質としては、水素も酸素も共に標準海水(standard mean ocean water: SMOW)が用いられる。水素の安定同位体は1Hと2Hの2種類で、1HはH、2HはD(重水素deuteriumの頭文字)、水素安定同位体比はδDと表記されることが多い。酸素の安定同位体は16O、17Oおよび18Oの3種類であるが、17Oは存在比が低いため、酸素の安定同位体比としては16O/18Oが使われる。水素および酸素についてそれぞれRをD/H、16O/18Oとすると、水素安定同位体比δDおよび酸素安定同位体比δ18Oは次のように定義される。
δD, δ18O(‰)=(Rサンプル/R標準物質 - 1)×1000;
水素安定同位体比は非常に変動が大きく、自然環境下で700‰にもなる。雨水の水素安定同位体比は-10~-400‰である。雨水の酸素安定同位体比は水素安定同位体比と直線的な相関関係があり
δD=8×δ18O + 10 (‰)
とあらわされる。
水素・酸素安定同位体比の決定要因
水が蒸発するときには重い安定同位体が分別されるため、水蒸気には軽い安定同位体(16O、H)が多く、残った水には重い安定同位体(18O、D)が多くなる。逆に水蒸気が凝固して水滴になるときには、軽い安定同位体が分別される。蒸発と凝固による同位体分別の結果、気温と降雨の安定同位体比との間には正の相関関係が成立し、気温が高いほど降水のδ18Oはプラスになる。葉の水のδDおよびδ18Oは、供給源の水のδDおよびδ18Oと、蒸散の影響を受ける。このことから、炭素安定同位体比δ13Cと同様に、植物の基質である水の起源をδDおよびδ18Oから推定することが可能である。また、植物の水利用と関連させることも可能であると考えられるが、蒸散と同位体分別との関係モデル(Craig-Gordonモデル)は実測値とのずれが大きく未だに不完全であり、現在も修正が試みられている。